Antibodies (immunoglobulin)
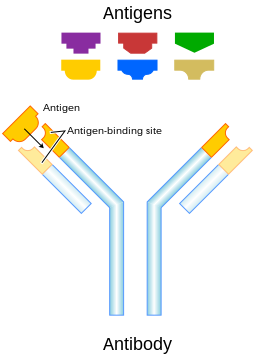
Each antibody binds to a specific antigen; an interaction similar to a lock and key
An antibody (Ab) or called immunoglobulin (Ig), is a large Y-shaped protein produced by B-cells that the immune system uses to identify and neutralize foreign objects. The antibody recognizes an antigen which is a unique part of the foreign target. Each tip of the "Y" of an antibody contains a paratope that is specific for one particular epitope (similarly analogous to a key) on an antigen, allowing these two structures to bind together with precision. Using this binding mechanism, an antibody can tag a microbe or an infected cell for attack by other parts of the immune system, or can neutralize its target directly.The production of antibodies is the main function of the humoral immune system.
Antibodies are secreted by a type of white blood cell called a plasma cell. Antibodies can occur in two physical forms, a soluble form that is secreted from the cell, and a membrane-bound form that is attached to the surface of a B cell and is referred to as the B cell receptor (BCR). The BCR is only found on the surface of B cells and facilitates the activation of these cells and their subsequent differentiation into either antibody factories called plasma cells, or memory B cells that will survive in the body and remember that same antigen so the B cells can respond faster upon future exposure. In most cases, interaction of the B cell with a T helper cell is necessary to produce full activation of the B cell and, therefore, antibody generation following antigen binding. Soluble antibodies are released into the blood and tissue fluids, as well as many secretions to continue to survey for invading microorganisms.
Antibodies are glycoproteins belonging to the immunoglobulin superfamily; the terms antibody and immunoglobulin are often used interchangeably. Antibodies are typically made of basic structural units each with two large heavy chains and two small light chains. There are several different types of antibody heavy chains, and several different kinds of antibodies, which are grouped into different isotypes based on which heavy chain they possess. Five different antibody isotypes are known in mammals, which perform different roles, and help direct the appropriate immune response for each different type of foreign object they encounter.
Though the general structure of all antibodies is very similar, a small region at the tip of the protein is extremely variable, allowing millions of antibodies with slightly different tip structures, or antigen binding sites, to exist. This region is known as the hypervariable region. Each of these variants can bind to a different antigen. This enormous diversity of antibodies allows the immune system to recognize an equally wide variety of antigens. The large and diverse population of antibodies is generated by random combinations of a set of gene segments that encode different antigen binding sites (or paratopes), followed by random mutations in this area of the antibody gene, which create further diversity. Antibody genes also re-organize in a process called class switching that changes the base of the heavy chain to another, creating a different isotype of the antibody that retains the antigen specific variable region. This allows a single antibody to be used by several different parts of the immune system.
History of Antibodies:
The first use of the term "antibody" occurred in a text by Paul Ehrlich. The term Antikörper (the German word for antibody) appears in the conclusion of his article "Experimental Studies on Immunity", published in October 1891, which states that "if two substances give rise to two different antikörper, then they themselves must be different". However, the term was not accepted immediately and several other terms for antibody were proposed; these included Immunkörper, Amboceptor, Zwischenkörper, substance sensibilisatrice, copula, Desmon, philocytase, fixateur, and Immunisin. The word antibody has formal analogy to the word antitoxin and a similar concept to Immunkörper.The study of antibodies began in 1890 when Kitasato Shibasaburō described antibody activity against diphtheria and tetanus toxins. Kitasato put forward the theory of humoral immunity, proposing that a mediator in serum could react with a foreign antigen. His idea prompted Paul Ehrlich to propose the side chain theory for antibody and antigen interaction in 1897, when he hypothesized that receptors (described as “side chains”) on the surface of cells could bind specifically to toxins – in a "lock-and-key" interaction – and that this binding reaction was the trigger for the production of antibodies. Other researchers believed that antibodies existed freely in the blood and, in 1904, Almroth Wright suggested that soluble antibodies coated bacteria to label them for phagocytosis and killing; a process that he named opsoninization.

Michael Heidelberger
In the 1920s, Michael Heidelberger and Oswald Avery observed that antigens could be precipitated by antibodies and went on to show that antibodies were made of protein. The biochemical properties of antigen-antibody binding interactions were examined in more detail in the late 1930s by John Marrack. The next major advance was in the 1940s, when Linus Pauling confirmed the lock-and-key theory proposed by Ehrlich by showing that the interactions between antibodies and antigens depended more on their shape than their chemical composition. In 1948, Astrid Fagreaus discovered that B cells, in the form of plasma cells, were responsible for generating antibodies.
Further work concentrated on characterizing the structures of the antibody proteins. A major advance in these structural studies was the discovery in the early 1960s by Gerald Edelman and Joseph Gally of the antibody light chain, and their realization that this protein was the same as the Bence-Jones protein described in 1845 by Henry Bence Jones. Edelman went on to discover that antibodies are composed of disulfide bond-linked heavy and light chains. Around the same time, antibody-binding (Fab) and antibody tail (Fc) regions of IgG were characterized by Rodney Porter.Together, these scientists deduced the structure and complete amino acid sequence of IgG, a feat for which they were jointly awarded the 1972 Nobel Prize in Physiology or Medicine. The Fv fragment was prepared and characterized by David Givol. While most of these early studies focused on IgM and IgG, other immunoglobulin isotypes were identified in the 1960s: Thomas Tomasi discovered secretory antibody (IgA) and David S. Rowe and John L. Fahey identified IgD, and IgE was identified by Kimishige Ishizaka and Teruko Ishizaka as a class of antibodies involved in allergic reactions. In a landmark series of experiments beginning in 1976, Susumu Tonegawa showed that genetic material can rearrange itself to form the vast array of available antibodies.
Forms of Antibodies:
The membrane-bound form of an antibody may be called a surface immunoglobulin (sIg) or a membrane immunoglobulin (mIg). It is part of the B cell receptor (BCR), which allows a B cell to detect when a specific antigen is present in the body and triggers B cell activation. The BCR is composed of surface-bound IgD or IgM antibodies and associated Ig-α and Ig-β heterodimers, which are capable of signal transduction.A typical human B cell will have 50,000 to 100,000 antibodies bound to its surface. Upon antigen binding, they cluster in large patches, which can exceed 1 micrometer in diameter, on lipid rafts that isolate the BCRs from most other cell signaling receptors. These patches may improve the efficiency of the cellular immune response. In humans, the cell surface is bare around the B cell receptors for several hundred nanometers, which further isolates the BCRs from competing influences.
Isotypes of Antibodies:
Antibodies can come in different varieties known as isotypes or classes. In placental mammals there are five antibody isotypes known as IgA, IgD, IgE, IgG and IgM. They are each named with an "Ig" prefix that stands for immunoglobulin, another name for antibody, and differ in their biological properties, functional locations and ability to deal with different antigens, as depicted in the table.
Name
|
Types
|
Description
|
Found in mucosal areas, such as the gut, respiratory tract and urogenital tract, and prevents colonization by pathogens.Also found in saliva, tears, and breast milk.
| ||
Functions mainly as an antigen receptor on B cells that have not been exposed to antigens. It has been shown to activate basophils and mast cells to produce antimicrobial factors.
| ||
Binds to allergens and triggers histamine release from mast cells and basophils, and is involved in allergy. Also protects against parasitic worms.
| ||
In its four forms, provides the majority of antibody-based immunity against invading pathogens. The only antibody capable of crossing the placenta to give passive immunity to the fetus.
| ||
Expressed on the surface of B cells (monomer) and in a secreted form (pentamer) with very high avidity. Eliminates pathogens in the early stages of B cell mediated (humoral) immunity before there is sufficient IgG.
|
Antibody Complexes

The antibody isotype of a B cell changes during cell development and activation. Immature B cells, which have never been exposed to an antigen, are known as naïve B cells and express only the IgM isotype in a cell surface bound form. B cells begin to express both IgM and IgD when they reach maturity
the co-expression of both these immunoglobulin isotypes renders the B cell 'mature' and ready to respond to antigen.
B cell activation follows engagement of the cell bound antibody molecule with an antigen, causing the cell to divide and differentiate into an antibody producing cell called a plasma cell. In this activated form, the B cell starts to produce antibody in a secreted form rather than a membrane-bound form. Some daughter cells of the activated B cells undergo isotype switching, a mechanism that causes the production of antibodies to change from IgM or IgD to the other antibody isotypes, IgE, IgA or IgG, that have defined roles in the immune system.
Structure:
Antibodies are heavy (~150 kDa) globular plasma proteins. They have sugar chains added to some of their amino acid residues. In other words, antibodies are glycoproteins. The basic functional unit of each antibody is an immunoglobulin (Ig) monomer (containing only one Ig unit); secreted antibodies can also be dimeric with two Ig units as with IgA, tetrameric with four Ig units like teleost fish IgM, or pentameric with five Ig units, like mammalian IgM.The variable parts of an antibody are its V regions, and the constant part is its C region.
- Immunoglobulin domains:
The Ig monomer is a "Y"-shaped molecule that consists of four polypeptide chains; two identical heavy chains and two identical light chains connected by disulfide bonds. Each chain is composed of structural domains called immunoglobulin domains. These domains contain about 70-110 amino acids and are classified into different categories (for example, variable or IgV, and constant or IgC) according to their size and function. They have a characteristic immunoglobulin fold in which two beta sheets create a “sandwich” shape, held together by interactions between conserved cysteines and other charged amino acids.
Several immunoglobulin domains make up the two heavy chains (red and blue) and the two light chains (green and yellow) of an antibody. The immunoglobulin domains are composed of between 7 (for constant domains) and 9 (for variable domains) β-strands
- Heavy chain:
There are five types of mammalian Ig heavy chain denoted by the Greek letters: α, δ, ε, γ, and μ.The type of heavy chain present defines the class of antibody; these chains are found in IgA, IgD, IgE, IgG, and IgM antibodies, respectively. Distinct heavy chains differ in size and composition; α and γ contain approximately 450 amino acids, while μ and ε have approximately 550 amino acids.In birds, the major serum antibody, also found in yolk, is called IgY. It is quite different from mammalian IgG. However, in some older literature and even on some commercial life sciences product websites it is still called "IgG", which is incorrect and can be confusing.
Each heavy chain has two regions, the constant region and the variable region. The constant region is identical in all antibodies of the same isotype, but differs in antibodies of different isotypes. Heavy chains γ, α and δ have a constant region composed of three tandem (in a line) Ig domains, and a hinge region for added flexibility; heavy chains μ and ε have a constant region composed of four immunoglobulin domains. The variable region of the heavy chain differs in antibodies produced by different B cells, but is the same for all antibodies produced by a single B cell or B cell clone. The variable region of each heavy chain is approximately 110 amino acids long and is composed of a single Ig domain.

1. Fab region
2. Fc region
3. Heavy chain (blue) with one variable (VH) domain followed by a constant domain (CH1), a hinge region, and two more constant (CH2 and CH3) domains.
4. Light chain (green) with one variable (VL) and one constant (CL) domain
5. Antigen binding site (paratope)
6. Hinge regions.
- Light chain:
In mammals there are two types of immunoglobulin light chain, which are called lambda (λ) and kappa (κ).A light chain has two successive domains: one constant domain and one variable domain. The approximate length of a light chain is 211 to 217 amino acids. Each antibody contains two light chains that are always identical; only one type of light chain, κ or λ, is present per antibody in mammals. Other types of light chains, such as the iota (ι) chain, are found in other vertebrates like sharks (Chondrichthyes) and bony fishes (Teleostei).
- CDRs, Fv, Fab and Fc Regions:
Some parts of an antibody have the same functions. The arms of the Y, for example, contain the sites that can bind two antigens (in general, identical) and, therefore, recognize specific foreign objects. This region of the antibody is called the Fab (fragment, antigen binding) region. It is composed of one constant and one variable domain from each heavy and light chain of the antibody.The paratope is shaped at the amino terminal end of the antibody monomer by the variable domains from the heavy and light chains. The variable domain is also referred to as the FV region and is the most important region for binding to antigens.More specifically, variable loops of β-strands, three each on the light (VL) and heavy (VH) chains are responsible for binding to the antigen. These loops are referred to as the complementarity determining regions (CDRs). The structures of these CDRs have been clustered and classified by Chothia et al. and more recently by North et al. In the framework of the immune network theory, CDRs are also called idiotypes. According to immune network theory, the adaptive immune system is regulated by interactions between idiotypes.
The base of the Y plays a role in modulating immune cell activity. This region is called the Fc (Fragment, crystallizable) region, and is composed of two heavy chains that contribute two or three constant domains depending on the class of the antibody.Thus, the Fc region ensures that each antibody generates an appropriate immune response for a given antigen, by binding to a specific class of Fc receptors, and other immune molecules, such as complement proteins. By doing this, it mediates different physiological effects including recognition of opsonized particles, lysis of cells, and degranulation of mast cells, basophils and eosinophils.
Structure prediction:
The importance of antibodies in health care and the biotechnology industry demands knowledge of their structures at high resolution. This information is used for protein engineering, modifying the antigen binding affinity, and identifying an epitope, of a given antibody. X-ray crystallography is one commonly used method for determining antibody structures. However, crystallizing an antibody is often laborious and time consuming. Computational approaches provide a cheaper and faster alternative to crystallography, but their results are more equivocal since they do not produce empirical structures. Online web servers such as Web Antibody Modeling (WAM) and Prediction of Immunoglobulin Structure (PIGS) enables computational modeling of antibody variable regions. Rosetta Antibody is a novel antibody FV region structure prediction server, which incorporates sophisticated techniques to minimize CDR loops and optimize the relative orientation of the light and heavy chains, as well as homology models that predict successful docking of antibodies with their unique antigen.REFERENCE:
http://en.wikipedia.org/wiki/Antibody






0 comments:
Post a Comment